лекция 6.
Лекция 6.
Генетические основы онтогенеза. Генетика иммунитета
План
1. ОБЩИЕ ПОНЯТИЯ
2. ДИФФЕРЕНЦИАЛЬНАЯ АКТИВНОСТЬ ГЕНОВ - ОСНОВА КЛЕТОЧНОЙ ДИФФЕРЕНЦИРОВКИ. РОЛЬ ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ НА РАННИХ ЭТАПАХ РАЗВИТИЯ
3. КРИТИЧЕСКИЕ ПЕРИОДЫ РАЗВИТИЯ
4. ОСОБЕННОСТИ РАЗВИТИЯ ПРОКАРИОТ И ЭУКАРИОТ
5. ПЕНЕТРАНТНОСТЬ И ЭКСПРЕССИВНОСТЬ ГЕНОВ
6. ПЛЕЙОТРОПНОЕ ДЕЙСТВИЕ ГЕНА
7. ВЗАИМОДЕЙСТВИЕ ГЕНОТИПА И СРЕДЫ
ОБЩИЕ ПОНЯТИЯ
1. Онтогенез — непрерывный процесс количественных и качественных изменений, происходящих в организме в течение всей жизни при постоянном взаимодействии генотипа и условий среды.
2. Термины «онтогенез» и «филогенез» ввел в биологию немецкий зоолог Е. Геккель. Он же сформулировал и обосновал (1866) биогенетический закон. Термин «онтогенез» означает процесс индивидуального развития особи, «филогенез* — история развития вида. Согласно биогенетическому закону индивидуальное развитие особи (онтогенез) является как бы кратким повторением филогенеза.
3. Позднее А. Северцев уточнил и дополнил положения биогенетического закона. Он доказал, что в процессе онтогенеза происходит выпадение отдельных этапов исторического развития, повторение зародышевых стадий предков, а не взрослых форм, возникновение изменений, мутаций, которых не было у предков. Полезные мутации передаются по наследству и включаются в филогенез (например, сокращение числа позвонков у бесхвостых земноводных), вредные - ведут к гибели зародыша. Таким образом, онтогенез не только повторяет филогенез, но и является источником новых направлений филогенеза. Онтогенез представляет собой цепь сложных последовательных морфологических, биохимических и функциональных преобразований, происходящих в организме с начала развития до смерти.
4. Онтогенетика изучает генетические основы индивидуального развития путем изучения действия генов на обменные и морфологические процессы в системе онтогенеза.
5. Онтогенез животных включает два основных взаимосвязанных процесса — рост и развитие. Под ростом понимают процесс увеличения размеров организма, его массы, происходящий за счет накопления в нем активных, главным образом белковых, веществ. В основе роста лежит увеличение числа и размеров клеток и неклеточных образований. Под развитием понимают качественные изменения — процессы усложнения структуры организма, специализацию, дифференциацию и интеграцию его органов и тканей.
6. Одна из основных проблем биологии — выяснение вопроса: каким образом из одной-единственной клетки возникает множество разнообразных типов клеток, значительно различающихся между собой строением, функцией, и как в процессе онтогенеза идет становление признаков и свойств организма? Из однойединственной клетки - оплодотворенного яйца - возникает много различных типов клеток, образующих тела многоклеточных организмов, структура зиготы со всеми ее особенностями, т.е. ее генотип, предопределяет в определенной степени конечный результат развития, но действительный результат зависит и от условий развития. В зиготе заложены все возможности для развития в определенных условиях среды целостного организма со свойствами, характерными для представителей данного вида, хотя в себе самой она не содержит всех признаков и особенностей взрослого организма. Она обладает способностью повторять в общих чертах путь развития родителей и более далеких предков. Однако повторение этого пути никогда не бывает абсолютным, т.к. сами половые клетки ни-когда не бывают полностью тождественными, а условия внешней среды, в которой происходит развитие, всегда чем-либо различаются. К тому же обладающая многими наследственными потенциями зигота не реализует их целиком: развиваются лишь те из них, для которых имеются соответствующие условия. Развитие происходит в рамках наследственности.
7. Проблема изучения механизма генетического контроля онтогенеза важное теоретическое и практическое значение для успешного решения таких вопросов, как селекция животных и растений, профилактика и лечение генетически обусловленных болезней у животных и человека.
Связь между нормальным развитием, злокачественным перерождением, иммуногенезом, старением стала одной из причин интереса ученых к механизмам онтогенеза. Другая важная причина такого повышенного интереса - это успехи молекулярной биологии и генетики, которые создали стройную систему представлений о механизмах проявления генов и регуляции их действия. Эти успехи достигнуты в основном в исследованиях на бактериях и фагах. Для биологии характерно возрастающее стремление к исследованию аналогичных процессов у более высокоорганизованных существ. Но все же и в настоящее время многие очень важные закономерности онтогенеза в генетическом плане остаются недостаточно выясненными.
Накопленный к настоящему времени фактический материал, связанный с закономерностями онтогенеза, условно группируется в следующем порядке: 1) роль и значение генов в регуляции биосинтеза и его онтогенетические особенности; 2) регуляция активности отдельных генов; 3) возрастные изменения клеточных структур, связанные с изменением активности отдельных генов; 4) генетические основы формирования морфологических признаков; 5) влияние мутации, трансплантации на формирование морфологических признаков и свойств.
РОЛЬ ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ НА РАННИХ ЭТАПАХ РАЗВИТИЯ. ДИФФЕРЕНЦИАЛЬНАЯ АКТИВНОСТЬ ГЕНОВ - ОСНОВА КЛЕТОЧНОЙ ДИФФЕРЕНЦИРОВКИ.
Центральный вопрос генетики развития: каким образом из одного клеточного генотипа возникает много сотен различных клеточных фенотипов, из одного яйца возникает до тысячи различных типов клеток.
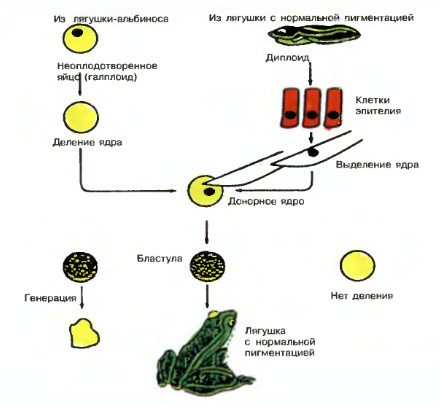
Выяснение механизмов дифференцировки клеток —одна из главных задач современной биологии. Поскольку дифференцировка необратима, некоторые ученые в конце прошлого века считали, что в ее основе лежит неравное распределение генов в те или иные дифференцирующиеся клетки в ходе последовательных клеточных делений. Это предположение было опровергнуто. В начале нашего века было показано, что каждая соматическая клетка имеет такой же набор хромосом, как и исходная оплодотворенная яйцеклетка.
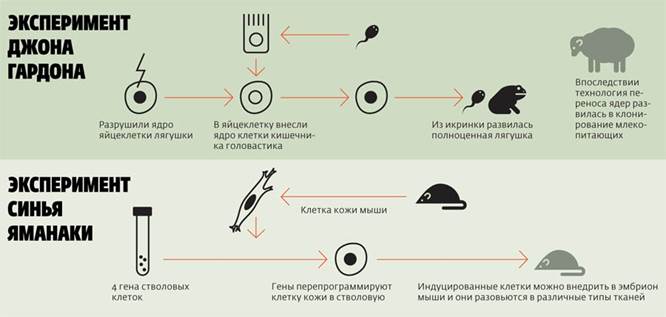
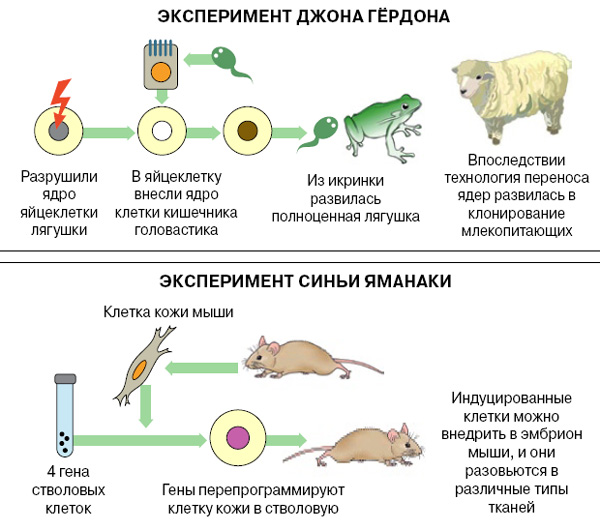
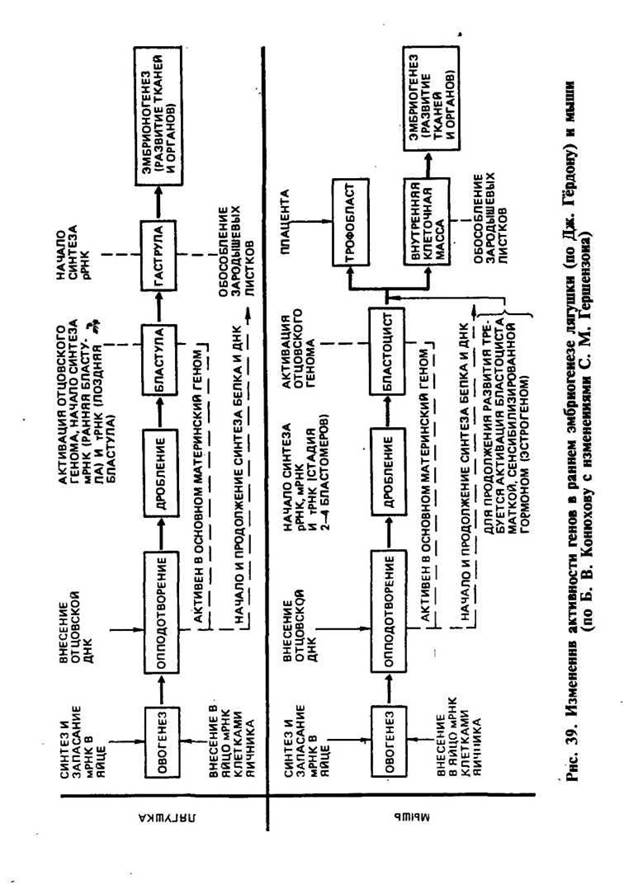
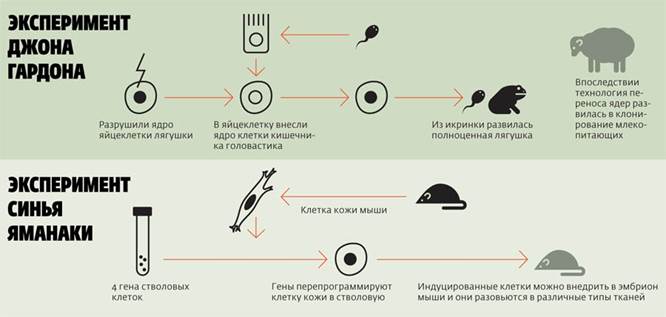
Доказательством являются специальные опыты по пересадке ядер. Дж. Гёрдон (1962) разрушал ядра яйцеклеток лягушки ультрафиолетовыми лучами и инъецировал в энуклеированные яйцеклетки ядра дифференцированных клеток кишечного эпителия плавающего головастика. Небольшой процент таких ядер обеспечивал развитие головастиков и нормальных лягушек. Опытами было показано, что ядра кишечных клеток содержат все гены, необходимые для дифференцировки всех типов клеток.
Это был один из первых успешных случаев искусственного клонирования животного, доказавший.
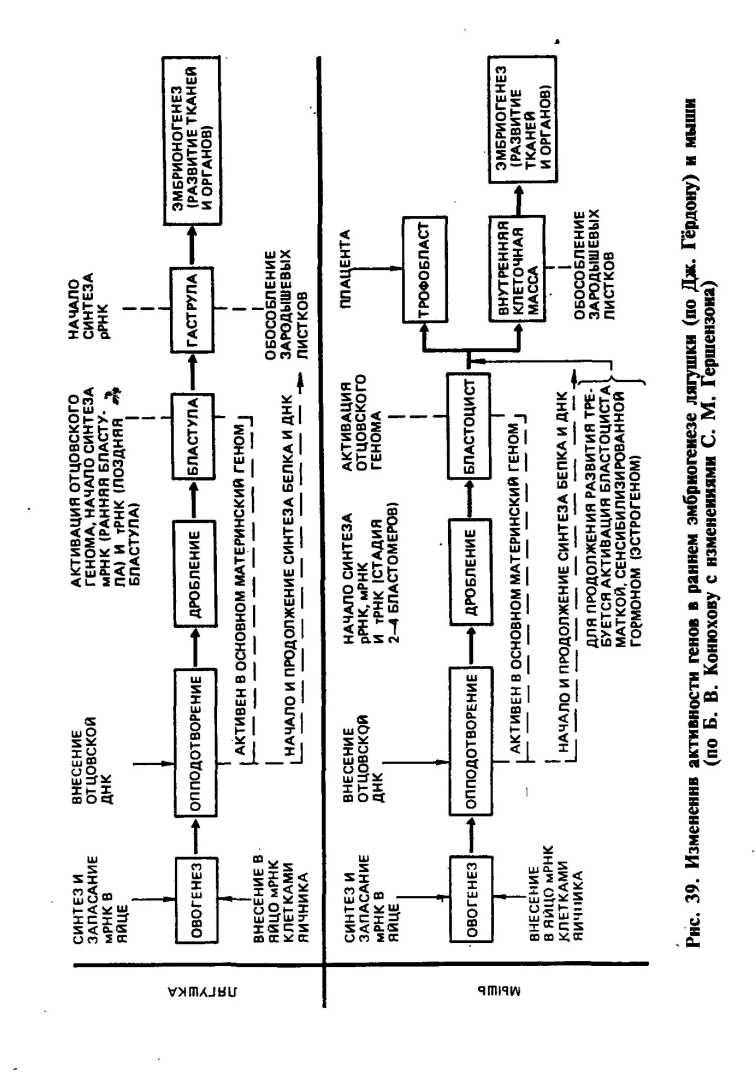
У животных в цитоплазме яйцеклетки до оплодотворения накапливается большое количество рибонуклеиновых кислот всех трех типов: иРНК, рРНК и тРНК. Они соединяются со специфическими белками-гистонами и образуют неактивные гранулы — инфорсомы. Через несколько минут после оплодотворения часть молекул иРНК инфорсом освобождается от белка, поступает на рибосомы цитоплазмы яйцеклетки и начинает синтез определенных белков, необходимых для начального развития зиготы. Начальный период развития зиготы проходит под контролем генов материнского организма, в частности иРНК яйцеклетки, обеспечивает синтез белков до стадии поздней бластулы. С начала стадии гаструляции и в дальнейших процессах онтогенеза синтез белка осуществляется под контролем ядерных генов обеих родительских форм. В эмбриогенезе лягушки синтез иРНК возобновляется после 10 делений дробления, когда зародыш состоит приблизительно из тысячи клеток.
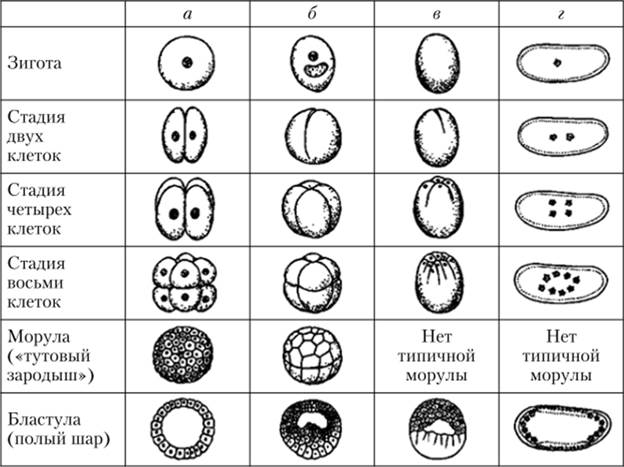
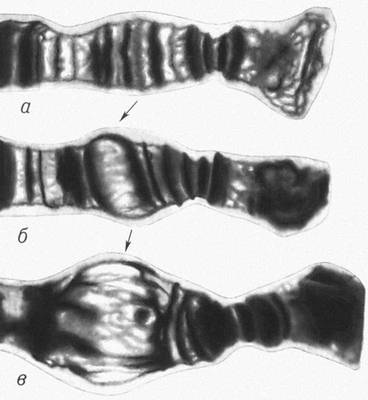
Дробление оплодотворенных яйцеклеток различных типов:
а — полное равномерное дробление у ланцетника; б — полное неравномерное дробление у лягушки; в — неполное дискоидальное дробление у птиц; г — неполное
поверхностное дробление у насекомых
В частности, в последующей работе по пересадке ядер Дж Гёрдон показал, что в течение первых десяти клеточных делений при развитии эмбриона лягушки в ядрах не наблюдается синтеза РНК. Клетки в этот период быстро делятся, реплицируют ДНК. Однако в клетках идет синтез белка. Дело в том, что у животных в период роста и созревания яйцеклетки в цитоплазме накапливается большое количество молекул РНК, которые, соединившись с белками-гистонами, образуют гранулы — информосомы.
Информосомы до оплодотворения яйцеклетки находятся в неактивном
На рисунке приведена схема изменения активности генов в раннем эмбриогенезе лягушки и мыши.
Одним из примеров дифференциальной активности генов в период органогенеза может служить процесс формирования пуффов в гигантских хромосомах дрозофилы. Гигантские хромосомы слюнных желез являются политенными и включают до 1000 нитей. Они имеют по длине определенный рисунок. На хромосомах видны диски, которые представляют собой соединение гомологичных генов. Было установлено, что на определенных стадиях отдельные диски деспирализуются и принимают форму вздутий, получивших название пуффов. В пуффах происходит интенсивный синтез молекул иРНК. Разные стадии развития личинок сопровождаются активностью определенных пуффов. Это говорит о том, что на разных этапах развития вступают в действие разные гены.
О неодновременной активности различных генов может свидетельствовать изменение состава белков организма в связи с возрастом. На стадиях раннего эмбрионального развития у человека идет образование гемоглобина F, который состоит из двух цепей полипептидов — α- и γ-цепей. Приблизительно с 13 нед эмбрионального развитии начинается синтез гемоглобина А, характерного для взрослого человека. У гемоглобина А цепь полипептида γ заменена на цепь β-цепей несколько иного строения. Цепь α у обоих гемоглобинов одинакова, и ее синтез контролируется одним и тем же геном. У новорожденного гемоглобин F составляет 70— 80 % общего количества. И только к году происходит полная замена гемоглобина F гемоглобином А. Полная замена гемоглобина F гемоглобином А у телят происходит к ПО—120-дневному возрасту.
Гемоглобин F обладает повышенным сродством к кислороду и позволяет сравнительно малому объёму крови плода выполнять кислородоснабжающие функции более эффективно. Однако гемоглобин F обладает меньшей стойкостью к разрушению и меньшей стабильностью в физиологически широком интервале pH и температур. В течение последнего триместра беременности и вскоре после рождения ребёнка гемоглобин F постепенно — в течение первых нескольких недель или месяцев жизни, параллельно увеличению объёма крови — замещается «взрослым» гемоглобином А (HbA), менее активным транспортёром кислорода, но более стойким к разрушению и более стабильным при различных значениях pH крови и температуры тела. Такое замещение происходит вследствие постепенного снижения продукции γ-цепей глобина и постепенного увеличения синтеза β-цепей созревающими эритроцитами. Первичная структура гемоглобина F отличается от первичной структуры гемоглобина А в тридцати девяти позициях (последовательность β-цепей по сравнению с таковой у γ-цепей). Эти отличия лежат в основе отличий физико-химических свойств гемоглобина А от гемоглобина F.
Обнаружены существенные возрастные изменения в количестве и составе белков сыворотки крови у телят в эмбриональный период. По данным В. М. Холода, первый период эмбрионального развития характеризуется низким содержанием сывороточных белков (2,62 г%), затем количество их постепенно с возрастом плода увеличивается и к 9 мес достигает 4,44 г%. Отношение альбуминов к глобулинам возрастает с 0,40 у 2-месячного плода до 1,21 к моменту рождения. В постэмбриональный период также наблюдаются изменения белкового спектра сыворотки крови. По данным А. С. Гурьяновой, у телок бурой латвийской порода содержание общего белка сыворотки крови с 3- до 18-месячного возраста увеличилось с 6,12 до 7,54 %, в том числе глобулинов с 3,03 до 4,24 %.
Некоторые органы и ткани специализируются на синтезе каких-то определенных белков, и количество РНК в них в отдельные периоды возрастает или снижается. И. Я. Шихов изучал содержание ДНК и РНК в вымени телок, нетелей и коров. Он обнаружил, что отношение количества РНК к количеству ДНК составляет в среднем у половозрелых телок 0,48, у нетелей и коров в конце стельности 1,0, у коров в начале лактации 2,34 (с большими колебаниями), в конце лактации 1,72. Наблюдалась высокая степень связи (г = 0,71) между содержанием РНК в вымени и удоем коров. Это показывает, что образование РНК усиливается, когда в вымени коров синтезируется много белка при высоких удоях, и снижается при уменьшении удоев.
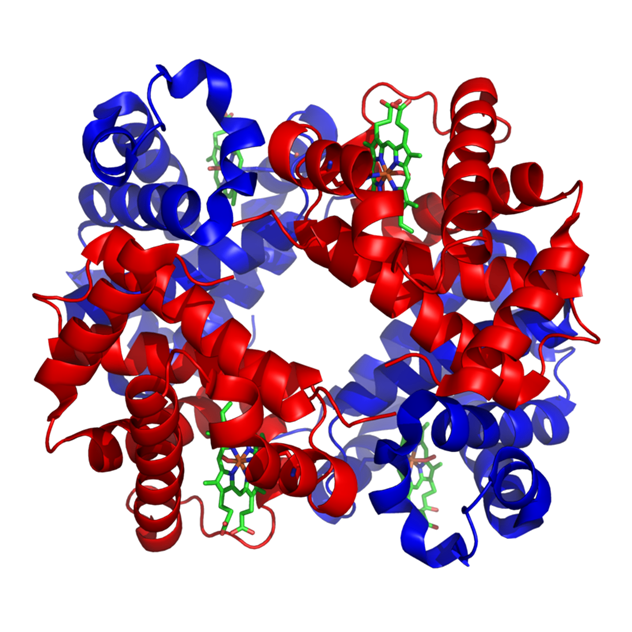
Структура человеческого гемоглобина. "'α"' и "'β"' субъединицы выделены красным и синим соответственно, а железосодержащий гем группы в зеленом цвете.
7.2. КРИТИЧЕСКИЕ ПЕРИОДЫ РАЗВИТИЯ
Генная система регуляции онтогенеза не всегда обеспечивает гармоничное развитие эмбриона из-за несоответствия самих условий развития возможностям генотипа.
Ученые-эмбриологи установили, что резкое изменение среды в определенные периоды эмбрионального развития организма может вызвать гибель зародыша или развитие уродств. Такие периоды принято называть критическими. Они обнаружены в онтогенезе млекопитающих, птиц, рептилий, амфибий и рыб. Критические периоды можно выявить после поздней бластулы, они предшествуют основным процессам морфогенеза. В этот период происходит развитие эмбриона под контролем генетической информации обеих родительских особей.
Наиболее изучены внешние факторы, влияющие в критические периоды на процесс онтогенеза у рыб и кур, несколько меньше — у млекопитающих и других классов. Так, у рыб нормальный онтогенез зависит от температуры воды и содержания в ней кислорода, причем у разных видов потребность в этих факторах различна: вьюн менее чувствителен к этим факторам, чем форель, лосось. У кур на эмбриогенез большое влияние оказывают температура и влажность воздуха в период инкубации. Эмбрионы особенно чувствительны к данным факторам на 2—3-и сутки инкубации, когда происходит образование системы кровообращения; на 8—9-е сутки — в период интенсивного морфогенеза; на 19-е сутки, когда происходит переход зародыша к легочному типу дыхания. Критические периоды онтогенеза определены у хомяков, морских свинок, кроликов и других животных. У крупного рогатого скота наблюдается повышение эмбриональной смертности в первые дни развития зиготы, что свидетельствует о критическом периоде.
Однако не следует путать критический период с чувствительным периодом, который проявляется тогда, когда действуют гены, контролирующие развитие данного признака.
По времени проявления чувствительный период опережает критический период, который наступает лишь тогда, когда повреждение в генах или генных продуктах цитоплазмы реализуется как уродство или деталь и не может компенсироваться в данных условиях активностью других генов.
ОСОБЕННОСТИ РАЗВИТИЯ ПРОКАРИОТ И ЭУКАРИОТ
У прокариот гены представляют собой непрерывную последовательность триплетов, обеспечивающих кодирование коллинеарной последовательности аминокислот в определенной белковой молекуле. У эукариот многие гены имеют участки, не несущие информации (интроны). Так, в гене, кодирующем альбумин крови крысы, содержится 13 интронов, в гене овальбумина курицы — 7 интронов.
Прокариоты вынуждены обеспечивать все необходимое для своего развития за счет активности почти всех имеющихся у них генов, организованы весьма просто, а различия их признаков проявляются прежде всего на молекулярном уровне (например, способность или неспособность к синтезу определенных соединений, к усвоению веществ и энергии развития клетки; чув-ствительность к антибиотику). Признаки прокариот можно назвать элементарными, ибо для них характерна простая связь: от гена к признаку.
У эукариот, в отличие от прокариот, происходит дифференцировка между разными клетками организма, что открывает широкие возможности для дистанционного (опосредованного) действия генов. Для многоклеточных эукариот характерно наличие хромосом, количество ДНК в каждой из которых превышает количество ДНК в геноме прокариот. Следует также отметить, что с появлением диплоидности (2л) у эукариот обозначилось значительно больше генных взаимодействий по сравнению с прокариотами. Существенное значение приобрели системы доминирования и супрессии, влияющие на баланс действия генов сложного многоклеточного организма.
Признак у эукариот, как правило, определяется не одним, а многими генами (полигенами). Например, на живую массу сельскохозяйственных животных оказывают влияние многие гены, основные из которых: а) гены, контролирующие ранние периоды развития в эмбриогенезе; б) гены дифференцировки органов и тканей; в) гены, увеличивающие живую массу в постэмбриональный период жизни; г) гены, влияющие на отложение подкожного жира и жира на внутренних органах.
Эукариоты: наличие дифференцированных органов и тканей, состоящих из узкоспециализированных клеток. В этих клетках в активном состоянии находится только та часть генетической информации, которая необходима для синтеза строго определенных белков. (например, гены, кодирующие синтез миозина в мышцах), и гены, ответственные за выполнение ограниченных функций, например за синтез гемоглобина крови, кератина волос и т. д.
На синтез белков у эукариот в процессе онтогенеза оказывают влияние: репликация ДНК, стабильность иРНК, каскадная и гормональная регуляция активности генов.
Интенсивность репликации ДНК зависит от дефференцировки клеток: в высокодефференцированных клетках (нейроны, мышечные клетки, клетки печени) репликация ДНК не происходит довольно длительное время, в отлие от клеток эпителия кишечника, костного мозга.
Стабильность иРНК. В отличие от прокариот иРНК у эукариот, особенно в клетках животных, относительно стабильна и может длительное время служить матрицей белкового синтеза, а также сохраняться в цитоплазме в виде инфорсом. Так, у человека длительность жизни ретикулоцитов до превращения их в эритроциты равна шести суткам. Ядра у них отсутствуют, но синтез специализированных молекул белка в них протекает на иРНК, образовавшихся в ядрах на предшествующей стадии нормобласта. По-видимому, для эукариот характерно иногда довольно продолжительное неодновременное протекание процессов транскрипции и трансляции. Таким образом, у высших организмов возможно образование безъядерных клеток, которые могут нормально функционировать за счет ранее синтезированных иРНК.
Каскадная регуляция активности генов. у эукариот что в клетке происходит одновременное включение или выключение большой группы генов, локализованных в разных молекулах ДНК, разных хромосомах. Эта регуляция осуществляется под воздействием специализированных весьма разнообразных сигнальных веществ, активно синтезируемых в клетках других тканей и поступающих в клетки данной ткани.
Гормональная регуляция активности генов. У высших животных важное значение имеет гормональная регуляция активности генов. Гормоны, вырабатываемые железами внутренней секреции, активируют синтез соответствующих белков. Они могут иметь белковую и небелковую природу, но синтез каждого из них осуществляется под генетическим контролем ДНК. Выделяясь из соответствующих желез в кровь, гормоны разносятся по всему организму, вступают в контакт с соответствующими клетками и активируют их гены;
Различные гормоны стимулируют активность генов у сельскохозяйственных животных, а следовательно, оказывают влияние на их продуктивность. Например, гормон роста соматотропин, вырабатываемый гипофизом индуцирует синтез белков во многих клетках одновременно. При введении соматотропина в организм телят повышается прирост их живой массы на 15—20 %, соматотропин увеличивает скорость роста индеек.
На активность соответствующих генов оказывают существенное влияние цитоплазма дифференцированных клеток и белки- гистоны. При дифференцировке клетка приобретает способность реагировать только на определенные раздражители, в результате чего она синтезирует только те белки, которые необходимы для ее дальнейшего функционирования.
Общая схема взаимодействия генов в онтогенезе у многоклеточных организмов (рис. 7.3) четко показывает, что каждый этап развития, ведущий к образованию зачатка новой ткани, зависит от взаимодействия генов данной клетки с генами других клеток. В результате такого действия происходит изменение цитоплазмы, что снимает репрессию и индуцирует активность тех генов, которые, в конечном счете, приводят к дифференцировке, а следовательно, к возникновению зачатия новой ткани или органа.
На молекулярном уровне такой процесс новообразования осуществляется путем транскрипции и трансляции, а также появлением в клетках качественно новых белков.
С
Схема взаимосвязей в системе генетического контроля индивидуального развития
7.2.1. ПЕНЕТРАНТНОСТЬ И ЭКСПРЕССИВНОСТЬ ГЕНОВ
Один и тот же признак может проявляться или не проявляться у особей родственных групп. Это явление называется пенетрантностью гена. Пенетрантность определяют по проценту особей в популяции, у которых данный ген проявился. Например, у мышей известна мутация изогнутости хвоста («поросячий хвост»), которая имеет неполную пенетрантность и проявляется у 16,7 % потомков.
Экспрессивность гена —степень фенотипического проявления гена как мера силы его действия, определяемая по степени развития самого признака.
Действие одних генов в онтогенезе может быть более или менее постоянным, стойким в своем проявлении или варьировать в зависимости от внешних условий. Рецессивные гены, которые в обычных условиях в гетерозиготном состоянии фенотипически не проявляются, могут проявиться при измененных условиях.
Классическим примером экспрессивности и различного фенотипического проявления гена может служить окраска меха у кролика, определяемая серией множественных аллелей гена С (СА, С”, Сл, Q.
Гималайская окраска меха определяется аллелем С* и фенотипически проявляется в белой окраске меха, но с черной окраской кончиков лап, ушей, носа и хвоста. Окраска меха зависит от биохимических реакций, протекающих в клетках кожи, контролирующих выработку меланина, и от температуры окружающей среды. Кролик, выращенный при температуре выше 30 °C, оказывается сплошь белым. Если выщипать небольшой участок белой шерсти и систематически его охлаждать, то на нем вырастет черная шерсть. Пониженная температура влияет на активность генов, контролирующих выработку определенных ферментов.
Важное значение для характера проявления генов имеют наличие и активность генов-модификаторов, определяющих степень экспрессивности генов в зависимости от специфических условий среды. В постэмбриональный период, когда идет процесс формирования конкретных признаков, присущих данной особи, генотип проявляется в фенотипе как система взаимосвязанных генов. В этой системе развитие одного признака может зависеть от взаимодействия многих генов и один ген может влиять на развитие и проявление нескольких признаков.
ПЛЕЙОТРОПНОЕ ДЕЙСТВИЕ ГЕНА
Приведем несколько классических примеров плейотропного действия генов:
1. Желтая мышь, у которой ген желтой окраски у гетерозигот обусловливает замедление обмена веществ и склонность к ожирению, некоторое увеличение размеров скелета, определенную устойчивость к заболеванию раком молочной железы.
2. У кур ген F вызывает курчавоперость. Перья у такой птицы обламываются, и со временем она теряет существенную часть перьевого покрова, что лишает ее возможности взлетать на насест. Одновременно у птицы в гомозиготном состоянии усиливается дыхание и потребление воды для поддержания оптимальной температуры тела, развивается гипертония сердечной мышцы, наблюдается задержка половой зрелости, что, в конечном итоге, приводит к снижению яйценоскости, оплодотворенности и выводимости яиц.
3. У норок большинство мутаций, вызывающих изменения окраски волосяного покрова, рецессивно и в силу плейотропии снижает их плодовитость и жизнеспособность. Ген пастелевой окраски приводит к уменьшению пигментированности оболочки глаз, когтей и губ, а также к синдрому закидывания головы у отдельных особей.

Плейотропное действие гена в большинстве случаев явление нежелательное для практической селекции растений и животных. Дело в том, что плейотропия затрудняет отбор, как правило, полезных признаков форм, так как положительное изменение одного признака может одновременно (чаще так и бывает) сопровождаться отрицательным изменением другого признака. Меланосаркомой чаще всего заболевают лошади серой масти.
Поэтому плейотропия указывает на связь процессов онтогенеза и взаимную связь генов в генной системе клетки и организма в целом.
ВЗАИМОДЕЙСТВИЕ ГЕНОТИПА И СРЕДЫ
Под взаимодействием «генотип—среда» понимается специфичность существования породы, линии или отдельной особи в разных условиях среды. Другими словами, одна среда лучше отвечает требованиям генотипа и способствует лучшему проявлению генетических задатков породы, линии, отдельной особи, в то время как в других условиях среды этого не происходит. Это положение наглядно проиллюстрировано на теоретическом примере (Д. Лэс- ли, 1982), в котором использованы три различных генотипа (А, В,С) и две среды (7, 2) (рис. 7.4). По генотипам А и Вне выявлено взаимодействия между генотипом и средой, поскольку животные этих генотипов имели сравнительно одинаковые (по направлению изменения) показатели продуктивности как в условиях

|
среды 7, так и в условиях среды 2, хотя генотип А по развитию признака был лучше, чем генотип В в обеих средах. В то же время наглядно показано взаимодействие «генотип—среда» по особи С. Так, генотип С превосходил по показателям продуктивности генотипы А и В в среде 7, но уступал им в условиях среды 2. Данный пример показывает, насколько важен првильный выбор линии или породы для конкретных условий среды, если мы стремимся получить оптимальную продуктивность.
Таким образом, реакция одного и того же организма особи на изменяющиеся условия внешней
среды будет далеко не одинаковой. Это лишний раз подтверждает, что наследуется не готовый признак, а реакция организма на условия жизни. В разных условиях один и тот же генотип будет проявляться поразному, так как развитие каждого признака особи контролируется определенными генами, проявляющимися только при определенных внешних факторах. Жживотное, имеющее высокую продуктивность в одних условиях среды, может иметь среднюю или даже худшую продуктивность в другой среде, и наоборот. Например, отдельные гибриды (бройлеры, несушки) дают более высокую продуктивность при клеточном, а другие — при напольном содержании, поэтому оценка их окажется в разных условиях неодинаковой.
Эффект взаимодействия генотипа и среды зависит от степени наследуемости признака. Признаки с низкой наследуемостью (половая зрелость, жизнеспособность, яйценоскость, оплодотворен- ность и др.) сильнее подвержены взаимодействию «генотип—среда». В то же время по таким признакам, как живая масса взрослых особей, содержание жира и белка в молоке, масса яиц, качество яиц, характеризующихся высокой степенью наследуемости, взаимодействия практически не наблюдается. По-видимому, проводить оценку животных по высоконаследуемым показателям достаточно по результатам испытания в одном стаде.
Во всех случаях, когда наблюдается достоверное взаимодействие «генотип—среда», следует руководствоваться только генотипической оценкой животных.
Сотрудниками кафедры генетики и разведения МГАВМиБ имени К. И. Скрябина в ОАО «Марьинская птицефабрика» (Московская область) изучено влияние взаимодействия генотипа и среды на организм яичных кур. Установлено, что яйценоскость и жизнеспособность яичных кур существенно различались в зависимости от генетического происхождения птицы (два кросса) и системы содержания (от четырех типов клеточного оборудования и двух режимов освещения). Дисперсионным анализом выявлено, что доля влияния взаимодействия генотипа и среды на яйценоскость несушек равна 0,84 (И. И. Кочиш, 2003).
Таким образом, проблема взаимодействия генотипа и среды имеет важное значение в селекции животных. Взаимодействие «генотип—среда» оказывает определенное влияние на эффективность отбора генетически лучших особей, программу отбора, характер и степень корреляции в оценке племенных особей, распределение животных по их генетическому достоинству в зависимости от условий жизни, приспособленность различных пород, линий и отдельных особей к экологическим условиям. С практической точки зрения взаимодействие генотипа и среды — существенный фактор, поэтому перспективные для селекционно-племенной работы производители должны быть оценены и отобраны именно в тех же условиях среды, где будет получено и использовано их потомство.
7.3. ВОЗРАСТНЫЕ ИЗМЕНЕНИЯ ПРИЗНАКОВ
Общеизвестно, что подвижность, высокая интенсивность роста и развития, активность процессов восстановления, акклиматизация и адаптация к новым условиям обитания происходят быстрее у молодых животных по сравнению с особями более старшего возраста. Также неодинакова реактивность и чувствительность организма к внешним факторам воздействия в разные периоды жизни. С возрастом характер этих процессов изменяется, наступают необратимые изменения в клетках и органах животного, составляющие в совокупности синдром старения. Доказано, что сроки наступления возрастных изменений в известной мере определены генетически и имеют видовую обусловленность.
С возрастом состав белков организма изменяется, их разнообразие соответствует числу генов, кодирующих белковую структуру. Однако следует также иметь в виду явление множественного аллелизма, ведущее к генетическому полиморфизму белков и ферментов, а также тот факт, что белки после их образования могут различным образом модифицироваться за счет включения в них радикалов (ацетильных, метильных и других групп).
Для животноводства важное значение приобретает биологическое и хозяйственное долголетие сельскохозяйственных животных. Биологическое долголетие — это длительность жизни животного, прерываемая естественной смертью. Хозяйственное долголетие — это длительность использования животного и способность его сохранять экономически выгодный уровень продуктивности и давать качественное потомство, то есть не утратившего способность к воспроизводству. Например, продолжительность жизни африканского страуса составляет 70 лет, а хозяйственного использования лишь 40, лошади — 67 и 20, крупного рогатого скота — 36 и 12, овцы —- 12 и 8, свиньи — 11 и 7, кролика — 7 и 3, курицы — 3 и 1,5 года соответственно.
Многочисленные исследования показали, что долголетие обусловлено наследственностью, а не только условиями жизни. Например, долголетие у коров айрширской породы в хозяйстве конного завода № 1 Московской области сопровождается сохранением высокой молочности и воспроизводительной функции. В среднем по стаду от каждой коровы получено по 7,5 теленка при что повышало индекс плодовитости. Некоторые коровы прожили по 17 лет и дали по 15 телят. В среднем пожизненный удой коров-долгожительниц составил 60 тыс. кг и более, и при выбытии удой был еще высок — 4400—4700 кг.
Исходя из вышеизложенного, можно сделать вывод, что повысить долголетие животных и тем самым улучшить экономический эффект животноводческой отрасли можно селекционной работой за счет соответствующего подбора.
Контрольные вопросы и задания.
1. Дайте характеристику биогенетическому закону онтогенеза.
2. Как изменяется функция генов в онтогенезе животных: экспрессия, пенетрантность?
3. Назовите критические периоды развития у животных разных видов.
4. Какое влияние оказывает гормональная и каскадная регуляция активности генов на синтез белков?
5 Приведите примеры плейотропного действия генов.
6. Опишите особенности развития прокариот и эукариот.
7. Расскажите о взаимодействии генотипа и среды.
8. Что понимают под биологическим и хозяйственным долголетием животных
О неодновременной активности различных генов может свидетельствовать изменение состава белков организма в связи с возрастом. На стадиях раннего эмбрионального развития у человека идет образование гемоглобина F, который состоит из двух цепей полипептидов — α- и γ-цепей. Приблизительно с 13 нед эмбрионального развитии начинается синтез гемоглобина А, характерного для взрослого человека. У гемоглобина А цепь полипептида γ заменена на цепь β-цепей несколько иного строения. Цепь α у обоих гемоглобинов одинакова, и ее синтез контролируется одним и тем же геном. У новорожденного гемоглобин F составляет 70— 80 % общего количества. И только к году происходит полная замена гемоглобина F гемоглобином А. Полная замена гемоглобина F гемоглобином А у телят происходит к ПО—120-дневному возрасту.
Гемоглобин F обладает повышенным сродством к кислороду и позволяет сравнительно малому объёму крови плода выполнять кислородоснабжающие функции более эффективно. Однако гемоглобин F обладает меньшей стойкостью к разрушению и меньшей стабильностью в физиологически широком интервале pH и температур. В течение последнего триместра беременности и вскоре после рождения ребёнка гемоглобин F постепенно — в течение первых нескольких недель или месяцев жизни, параллельно увеличению объёма крови — замещается «взрослым» гемоглобином А (HbA), менее активным транспортёром кислорода, но более стойким к разрушению и более стабильным при различных значениях pH крови и температуры тела. Такое замещение происходит вследствие постепенного снижения продукции γ-цепей глобина и постепенного увеличения синтеза β-цепей созревающими эритроцитами.
Обнаружены существенные возрастные изменения в количестве и составе белков сыворотки крови у телят в эмбриональный период. По данным В. М. Холода, первый период эмбрионального развития характеризуется низким содержанием сывороточных белков (2,62 г%), затем количество их постепенно с возрастом плода увеличивается и к 9 мес достигает 4,44 г%. Отношение альбуминов к глобулинам возрастает с 0,40 у 2-месячного плода до 1,21 к моменту рождения. В постэмбриональный период также наблюдаются изменения белкового спектра сыворотки крови. По данным А. С. Гурьяновой, у телок бурой латвийской порода содержание общего белка сыворотки крови с 3- до 18-месячного возраста увеличилось с 6,12 до 7,54 %, в том числе глобулинов с 3,03 до 4,24 %.
Некоторые органы и ткани специализируются на синтезе каких-то определенных белков, и количество РНК в них в отдельные периоды возрастает или снижается. И. Я. Шихов изучал содержание ДНК и РНК в вымени телок, нетелей и коров. Он обнаружил, что отношение количества РНК к количеству ДНК составляет в среднем у половозрелых телок 0,48, у нетелей и коров в конце стельности 1,0, у коров в начале лактации 2,34 (с большими колебаниями), в конце лактации 1,72. Наблюдалась высокая степень связи (г = 0,71) между содержанием РНК в вымени и удоем коров. Это показывает, что образование РНК усиливается, когда в вымени коров синтезируется много белка при высоких удоях, и снижается при уменьшении удоев.
Однако цитоплазма играет очень важную роль в реализации наследственной информации и формировании некоторых призна-ков организма. Известно, что основная часть цитоплазмы посту-пает в зиготу с яйцеклеткой. Цитоплазма яйцеклетки отличается от цитоплазмы соматических клеток большим разнообразием бел¬ков, РНК и других видов молекул, синтезированных Биогенезе.
У амфибий и рыб в цитоплазме в период созревания ооцитов и в яйцеклетках были обнаружены в большом количестве фрагменты ДНК, а у кур — свободная ДНК и вителлогенин. В период созревания яйцеклеток происходит интенсивное насыщение цитоплазмы ДНК, иРНК, рРНК, тРНК, а также другими компонентами. У дрозофилы 15 фолликулярных клеток, окружающих яйцеклетку, проникают в нее цитоплазматическими выростами и насыщают цитоплазму митохондриями, ДНК, РНК, белками и другими компонентами. Установлено, что отдельные участки цитоплазмы яйцеклетки могут содержать факторы, определяющие судьбу тех или иных дифференцирующихся клеток. В результате неравномерного распределения веществ в цитоплазме яйцеклетки при дроблении зиготы идет неравнозначное распределение веществ (РНК, белков и др.) в бластомеры. Поэтому их цитоплазма оказывается различной.
Таким образом, ядро зиготы, образовавшееся в результате сли¬яния материнского и отцовского ядер и объединения их генети¬ческой информации, в начальный период развития зародыша не оказывает влияния на дробление зиготы и образование бластулы. Эмбрион в этот период развивается за счет РНК и других компо¬нентов, находящихся в цитоплазме яйцеклетки.
В процессе развития организма наблюдается сложное взаимодействие ядра и цитоплаз¬мы, влияние цитоплазмы на формирование некоторых признаков. Это происходит за счет цитоплазматической наследственности, которая обусловлена наличием в цитоплазме органелл (например, митохондрий, пластид), имеющих собственную ДНК, а следова¬тельно, и собственные гены. Известно, что у млекопитающих 99 % ДНК находится в ядре клетки и около 1 % в цитоплазме. У птиц ДНК в цитоплазме значительно больше, однако роль ее в передаче генетической информации еще далеко не ясна. В целом же у животных и растений определяющую роль в наследственности играет ядро клетки.
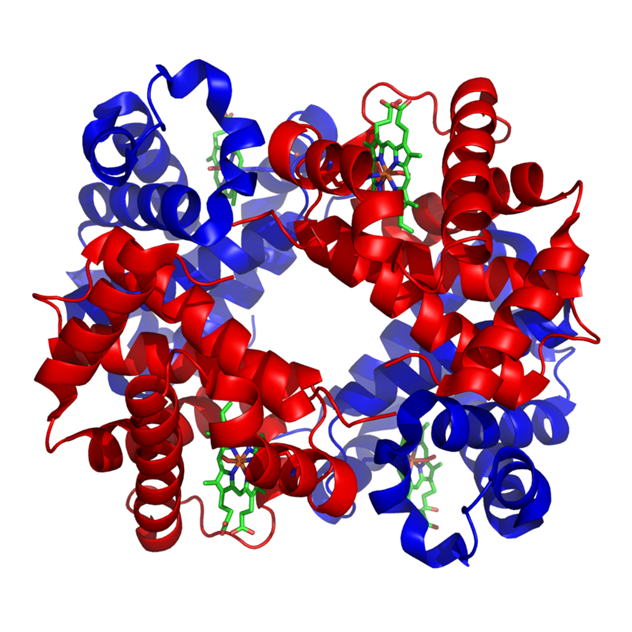
Структура человеческого гемоглобина. "'α"' и "'β"' субъединицы выделены красным и синим соответственно, а железосодержащий гем группы в зеленом цвете.
7.4. КРИТИЧЕСКИЕ ПЕРИОДЫ РАЗВИТИЯ
Генная система регуляции онтогенеза не всегда обеспечивает гармоничное развитие эмбриона из-за несоответствия самих условий развития возможностям генотипа.
Ученые-эмбриологи установили, что резкое изменение среды в определенные периоды эмбрионального развития организма может вызвать гибель зародыша или развитие уродств. Такие периоды принято называть критическими. Они обнаружены в онтогенезе млекопитающих, птиц, рептилий, амфибий и рыб. Критические периоды можно выявить после поздней бластулы, они предшествуют основным процессам морфогенеза. В этот период происходит развитие эмбриона под контролем генетической информации обеих родительских особей.
Наиболее изучены внешние факторы, влияющие в критические периоды на процесс онтогенеза у рыб и кур, несколько меньше — у млекопитающих и других классов. Так, у рыб нормальный онтогенез зависит от температуры воды и содержания в ней кислорода, причем у разных видов потребность в этих факторах различна: вьюн менее чувствителен к этим факторам, чем форель, лосось. У кур на эмбриогенез большое влияние оказывают температура и влажность воздуха в период инкубации. Эмбрионы особенно чувствительны к данным факторам на 2—3-и сутки инкубации, когда происходит образование системы кровообращения; на 8—9-е сутки — в период интенсивного морфогенеза; на 19-е сутки, когда происходит переход зародыша к легочному типу дыхания. Критические периоды онтогенеза определены у хомяков, морских свинок, кроликов и других животных. У крупного рогатого скота наблюдается повышение эмбриональной смертности в первые дни развития зиготы, что свидетельствует о критическом периоде.
Однако не следует путать критический период с чувствительным периодом, который проявляется тогда, когда действуют гены, контролирующие развитие данного признака.
По времени проявления чувствительный период опережает критический период, который наступает лишь тогда, когда повреждение в генах или генных продуктах цитоплазмы реализуется как уродство или деталь и не может компенсироваться
Раскрытие сложной структуры гена
В соответствии с представлениями классической генетики долгое время считалось, что ген — неделимая единица функции, рекомбинации и мутирования. Гены представлялись как бусинки, механически соединенные каким-то материалом в хромосоме. Вопрос о пересмотре представлений о гене как неделимой единице впервые был поставлен в 1929—1930 гг. А. С. Серебровским с сотр. Они изучили мутации гена scute (скыот), влияющего на развитие щетинок на теле дрозофилы, и обнаружили явление ступенчатого аллелизма. Было выявлено 14 мутаций гена scute: SCi, SC2, SC3 и т. д. Мутации отличались друг от друга редукцией щетинок на определенных участках тела: в одном случае не было щетинок на голове и предгруди, в другом — на предгруди и груди и т. д. Мы уже знаем, что при скрещивании особей с разными аллельными генами у потомков имеет место доминирование одного из них. В случае же серии аллелей гена scute у гетерозиготных особей был обнаружен частичный возврат к норме. У гетерозиготных потомков не развивались только те щетинки, которых не было у обоих родителей. Щетинки развивались нормально, если изменение наблюдалось только в одном из аллельных генов. Если, например, мутация SQ вызывала редукцию щетинок ABC, а мутация SC2 — редукцию щетинок BCD, то у гетерозиготы отсутствовали щетинки В и С и развивались нормально А и D. При графическом изображении взаимодействия несколько пар аллеломорфов получается как бы лестница, ступенями которой служат отдельные аллели гена scute (скьют): SCi - ABC; SC2 - BCD; SC3 - CDE.
Это явление получило название ступенчатого аллеломорфизма. Было показано, что ген не является единицей мутации, он дробим и имеет сложную структуру.
На основании проведенных исследований была сформулирована центровая теория гена. Согласно ей ген состоит из отдельных функциональных участков — центров, которые могут независимо изменяться при мутациях.
Глубокие исследования тонкой структуры генов фага Т4, поражающего кишечную палочку, были проведены американским генетиком С. Бензером. Дикий тип фага Т4 лизирует разные штаммы кишечной палочки, в том числе штаммы В и К. При этом в бактериальных культурах наблюдается появление мелких стерильных пятен. При посеве фага на штамм В были обнаружены и отобраны мутанты фага, названные rl 1 (г — от англ. rapid, lysis — быстрый лизис), образующие крупные стерильные пятна (по сравнению с фагом дикого типа) и не способные размножаться на штамме К (не образуют стерильных пятен). Для анализа выявленных мутантов С. Бензер разработал тест на компле-ментарность, который заключается в следующем: если бактерии штамма К заразить смесью двух разных мутантов rll и это не приведет к лизису клеток штамма К, значит,» мутации у обоих фагов затрагивают одну и ту же функцию и произошли в одном и том же гене. Если же при совместном заражении восстанавливается дикий тип, т. е. произойдет лизис клеток штамма К, значит, мутации комплементарны и произошли в разных генах. С. Бензер исследовал попарно очень большое число мутантов rll и установил, что они принадлежат к двум функциональным группам: А и В. Любой из мутантов группы А комплементарен любому из мутантов группы В, в то же время два мутанта, относящиеся к одной и той же группе А или В, некомплементарны, они затрагивают одну и ту же функциональную единицу, и в этом случае сохраняется мутантный фенотип. Наличие двух классов мутантов привело к выводу о существовании двух функциональных единиц в пределах rll генома фага Тф Каждая из этих единиц связана, очевидно, с синтезом специфического полипЙЬтида, необходимого для роста на штамме К. Генетическая единица функции, выявленная с помощью теста на компле-ментарность, была С. Бензером названа цистроном. В настоящее время термин «цистрон» используется как синоним гена.
Геном называется участок молекулы ДНК (у некоторых вирусов РНК), кодирующий первичную структуру полипептида, молекулы транспортной или рибосомной РНК или взаимодействующий с регу-ляторным белком.
Структурной единицей мутации и рекомбинации гена является одна пара нуклеотидов (или один нуклеотид в случае геномов, состоящих из одноцепочных ДНК или РНК). Для обозначения локализации мутаций в пределах гена применяется термин «сайт». Сайт может включать одну пару нуклеотидов, а в некоторых случаях, возможно, и больше. Размер генов в разных организмах различен, однако в среднем ген включает 1500 пар нуклеотидов. Самые короткие — гены, кодирующие тРНК. Они включают приблизительно 190 пар нуклеотидов. Но имеются и очень длинные гены. Например, ген фиброина шелка тутового шелкопряда включает 16 тыс. пар нуклеотидов. Таким образом, ген — очень сложная структура.
Установлено, что молекулярное строение генов эукариот отличается от генов прокариот. У последних гены представляют собой непрерывную последовательность триплетов, обеспечивающих кодирование колинеарной последовательности аминокислот в определенной полипептидной цепи. У эукариот многие гены имеют мозаичную структуру. Оказалось, что они составлены из кодирующих участков — экзонов, разделенных некодирующими участками — интронами. Например, в генах тяжелой цепи иммуноглобулинов не менее пяти экзонов и четырех интронов, в гене овальбумина (яичного белка) восемь экзонов и семь интронов.
В последние годы у прокариот обнаружены перекрывающиеся гены. Так, у некоторых РНК-содержащих бактериофагов Е. coli (R17 и др.) считались установленными три гена — репликазы, белка оболочки и созревания вирусной частицы. Однако был обнаружен четвертый ген, кодирующий белок L, включающий 75 аминокислотных остатков и блокирующий лизис зараженной клетки. Места для этого гена на РНК не было. Слева у гена обнаружили кодон-инициатор (АУГ), а справа — терминатор (УАА). Между ними расположено 75 триплетов. Оказалось, что ген локализован частично в гене белка оболочки (47 нуклеотидов), 36 нуклеотидов — в межгенном интервале и 142 нуклео-тида расположены в гене РНК — репликазы. Перекрывающиеся гены обнаружены также в некоторых одноцепочных ДНК фагов Е. coli и в ДНК вируса млекопитающих SV 40. Таким образом, используются одни и те же последовательности нуклеотидов для кодирования разных белков.
В конце 40-х годов XX в. Б. Мак-Клинток на основе генетических экспериментов на кукурузе предсказала наличие в геноме организмов подвижных элементов — «прыгающих» генов. В конце 60-х годов у бактерий обнаружено два основных класса мобильных («прыгающих») генов, которые различались по длине и сложности организации: 1-й — инсерционные последовательности, или lS-элементы. Длина их около 1000 пар нуклеотидов. Они содержат только один ген, ответственный за их перемещение; 2-й — транспозоны с длиной 3000—20 000 пар нуклеотидов. Транспозоны кроме системы транспозиции содержат дополнительные гены, определяющие устойчивость к антибиотикам, различным токсическим соединениям. В 70-х годах изучены мобильные гены у дрозофилы, затем было выявлено, что у эукариот они составляют не менее 5—10 % их генетического материала. Мобильные гены вносят в геном факторы нестабильности и изменчивости, что может играть, по-видимому, существенную роль в эволюции.
Дифференциальная активность генов - основа клеточной дифференцировки
Основная проблема биологии развития - каким образом из одной-единственной клетки - зародыша - возникает много различных типов клеток? Если выразить ее в генетических терминах, она становится центральным вопросом генетики развития: каким образом из одного клеточного генотипа возникает много сотен различных клеточных фенотипов, из одного яйца возникает до тысячи различных типов клеток.
Выяснение механизмов дифференцировки клеток —одна из главных задач современной биологии. Поскольку дифференцировка необратима, некоторые ученые в конце прошлого века считали, что в ее основе лежит неравное распределение генов в те или иные дифференцирующиеся клетки в ходе последовательных клеточных делений. Это предположение было опровергнуто. В начале нашего века было показано, что каждая соматическая клетка имеет такой же набор хромосом, как и исходная оплодотворенная яйцеклетка.
Доказательством являются специальные опыты по пересадке ядер. Дж. Гёрдон (1962) разрушал ядра яйцеклеток лягушки ультрафиолетовыми лучами и инъецировал в энуклеированные яйцеклетки ядра дифференцированных клеток кишечного эпителия плавающего головастика. Небольшой процент таких ядер обеспечивал развитие головастиков и нормальных лягушек. Опытами было показано, что ядра кишечных клеток содержат все гены, необходимые для дифференцировки всех типов клеток.
Это был один из первых успешных случаев искусственного клонирования животного, доказавший, что в ДНК специализированной клетки сохраняется вся информация, необходимая для дифференцировки всех клеточных типов.
В последующей работе по пересадке ядер Дж Гёрдон показал, что в течение первых десяти клеточных делений при развитии эмбриона лягушки в ядрах не наблюдается синтеза РНК. Клетки в этот период быстро делятся, реплицируют ДНК. Однако в клетках идет синтез белка. Дело в том, что у животных в период роста и созревания яйцеклетки в цитоплазме накапливается большое количество молекул РНК, которые, соединившись с белками-гистонами, образуют гранулы — информосомы.
Информосомы до оплодотворения яйцеклетки находятся в неактивном состоянии. Сразу же после оплодотворения мРНК освобождается от белков-гистонов, поступает в рибосомы цитоплазмы яйцеклетки и начинается синтез определенных белков по программе материнской ДНК. Поэтому начальный период развития зиготы осуществляется под контролем генов материнского организма. С начала стадии гаструляции и в дальнейшем синтез белка осуществляется под влиянием мРНК, образующейся в ядрах клеток эмбриона, т. е. под контролем генов обеих родительских особей.
На рисунке приведена схема изменения активности генов в раннем эмбриогенезе лягушки и мыши.
Одним из примеров дифференциальной активности генов в период органогенеза может служить процесс формирования пуф-фов в гигантских хромосомах дрозофилы. Гигантские хромосомы слюнных желез являются политенными и включают до 1000 нитей. Они имеют по длине определенный рисунок. На хромосомах видны диски, которые представляют собой соединение гомологичных генов. Было установлено, что на определенных стадиях отдельные диски деспирализуются и принимают форму вздутий, получивших название пуффов. При помощи использования радиоактивного уридина было установлено, что в пуффах происходит интенсивный синтез молекул иРНК. Разные стадии развития личинок сопровождаются активностью определенных пуффов. Это говорит о том, что на разных этапах развития вступают в действие разные гены.